

Ned Z. Carp MD, FACS
- Associate Professor of Surgery, Jefferson Medical College, Philadelphia,
- Pennsylvania
- Chief, Division of General Surgery, Lankenau Hospital, Wynnewood,
- Pennsylvania
The respiratory center is located in the medulla and regulates respiration with input from sensory (mechanoreceptor and chemoreceptor) feedback loops symptoms night sweats buy aricept cheap. Molecular and physiological determinants of pulmonary developmental biology: a review 10 medications that cause memory loss discount aricept 10 mg with visa. Lung inflammation and fibrosis: an alveolar macrophage-centered perspective from the 1970s to 1980s cancer treatment 60 minutes 5 mg aricept sale. Explain how surfactant affects lung compliance treatment using drugs is called purchase aricept 5 mg line, and describe its importance in maintaining unequal alveolar volumes treatment nurse 10 mg aricept amex. Thus a negative pressure in the pleural space is a pressure that is lower than atmospheric pressure treatment xeroderma pigmentosum 10mg aricept mastercard. Also in accordance with convention, pressures across surfaces such as the lungs or chest wall have been defined as the difference between the pressure inside and the pressure outside the surface. The pressure differences across the lung and across the chest wall are defined as the transmural (across a wall or surface) pressures. The mechanical properties of the lung and chest wall determine the ease or difficulty of this air movement. Lung mechanics is the study of the mechanical properties of the lung and chest wall (including the diaphragm, abdominal cavity, and anterior abdominal muscles). Lung mechanics is important for how the lungs work both normally and in the presence of disease, inasmuch as most lung diseases affect the mechanical properties of the lungs, chest wall, or both. In addition, death from lung disease is almost always due to respiratory muscle fatigue, which results from an inability of the respiratory muscles to overcome the altered mechanical properties of the lungs, chest wall, or both. Lung mechanics includes static mechanics (the mechanical properties of a lung whose volume is not changing with time) and dynamic mechanics (properties of a lung whose volume is changing with time). How a Pressure Gradient Is Created Air flows into and out of the lungs from areas of higher pres sure to areas of lower pressure. Before inspiration begins, the pleural pressure in normal individuals is approximately -3 to -5 cm H2O. Therefore, the pressure in the pleural space is negative in 447 Pressures in the Respiratory System In healthy people, the lungs and chest wall move together as a unit. Between these structures is the pleural space, which under normal conditions is best thought of as a potential (or virtual) space. Because the lungs and chest wall move together, changes in their respective volumes are equal during inspiration and exhalation. Volume changes in the lungs and chest wall are driven by changes in the surrounding pressure. This negative pressure is created by the inward elastic recoil pressure of the lung, and it acts to "pull the lung" away from the chest wall. The lung is not able, however, to pull away from the chest wall, inasmuch as the two function as a unit. Thus the inward elastic recoil pressure of the lung is balanced by the outward recoil of the chest wall. With the onset of inspiration, the muscles of the dia phragm and chest wall contract, which causes a downward movement of the diaphragm and an outward and upward movement of the rib cage. This negative pleural pressure is transmitted across the lung tissue and results in a decrease in alveolar pressure. As gas flows into the airways to the alveoli, the pressure gradient along the airways decreases, and flow stops when there is no longer a pressure gradient from atmospheric to alveolar pressure. The decrease in pleural pressure at the start of inspiration secondary to inspiratory muscle contraction is greater than the transmitted fall in alveolar pressure, and, as a result, transpulmonary pressure at the start of inspiration is positive (see Eq. Positive transpulmonary pressure is necessary to increase lung volume, and lung volume increases with increasing transpulmonary pressure. Because pleural pressure is negative in relation to atmospheric pressure during quiet breathing, the transmural pressure across the chest wall is negative (see Eq. On exhalation, the diaphragm moves higher into the chest, pleural pressure increases. In the alveoli, the driving force for exhalation is the sum of the elastic recoil of the lungs and pleural pressure (see Chapter 22). This relation ship between changes in pressure, changes in airflow, and changes in volume during inspiration and exhalation is displayed in. During tidal volume breathing in normal individuals, the decrease in alveolar pressure at the start of inspiration is small (1 to 3 cm H2O). It is much larger in individuals with airway obstruction because of the larger pressure drop that occurs across obstructed airways. Airflow stops in the absence of a pressure gradient, which occurs whenever alveolar pressure and atmospheric pressure are equal. Positive(inrelationtoatmospheric)pressuresarerepresentedabove the horizontal dotted line, and negative pressures are represented belowit. The patient then inhales maximally and exhales forcefully and completely, and the volume of exhaled air is measured. Both methods are used clinically and provide valuable information about lung function and lung disease. The helium dilution technique is the older and simpler method, but it is often less accurate than body plethysmography, which requires sophisticated and expensive equipment. C1, known concentration of an inert gas; C2, new (previously unknown) concentrationofthegas;V1,knownvolumeofabox;V2,lungvolume (initiallyunknown). The answer lies in the properties of the lung paren chyma and in the interaction between the lungs and the chest wall. The lung contains elastic fibers that (1) stretch when stress is applied, which results in an increase in lung volume, and (2) recoil passively when this stress is released, which results in a decrease in lung volume. Similarly, chest wall volume can increase when the respiratory muscles are stretched and decrease when respiratory muscle length is shortened. Decreasing lung volume results in shortening of the expira tory muscles, which, in turn, results in a decrease in muscle force. The decrease in lung volume is also associated with an increase in the outward recoil pressure of the chest wall. Pressure-Volume Relationships A number of important observations can be made from an examination of the pressurevolume curves of the lung, chest wall, and respiratory system. The transmural pressure for a healthy lung alone flattens at pressures higher than 20 cm H2O because the elastic limits of the lung have been reached. Thus further increases in transmural pressure produce little change in volume, and compliance (see the section "Lung Compliance") is low. Further distention is limited by the connective tissue (collagen, elastin) of the lung. If further pressure is applied, the alveoli near the lung surface can rupture, and air can escape into the pleural space. In a pneumothorax or when the chest is opened, as during thoracic surgery, the lungs and chest wall no longer function as a single unit. The lungs recoil until transpulmonary pressure is 0; the chest wall then increases in size until trans chest wall pressure is 0. The relationship between transpulmonary pressure and pleural, alveolar, and elastic recoil pressures is depicted in. Alveolar pressure is the sum of the pleural pres sure and elastic recoil pressure (Pel) of the lung: Equation 21. Graphically, lung compliance is the slope of the line between any two points on the deflation limb of the pressurevolume loop. For this reason, compliance is corrected for the lung volume at which it is measured (specific compliance;. Compliance is not often measured for clinical purposes because it requires placement of an esophageal balloon. The esophageal balloon, which is connected to a pressure transducer, is an excellent surrogate marker for pleural pressure, which is very difficult to measure directly. Surface tension is a force caused by water molecules at the airliquid interface that tends to minimize surface area, which makes inflating the lungs more difficult. The effect of surface tension on lung infla tion is illustrated by a comparison of the volumepressure curves of a salinefilled lung and of an airfilled lung. Higher pressure is necessary to fully inflate the lung with air than with saline because of the higher surface tension forces in airfilled lungs than in salinefilled lungs. Surface tension is a measure of the attractive force of the surface molecules per unit length of material to which they are attached. For a sphere (such as an alveolus), the relationship between the pressure within the sphere (Ps) and the tension in the wall is described by the law of Laplace: Equation 21. The alveoli are lined with a predominantly lipidbased substance called surfactant. Pulmonary surfactant serves several physiological roles, including (1) reducing the work of breathing by decreasing surface tension forces; (2) preventing collapse and sticking of alveoli on exhalation; and (3) stabilizing alveoli, especially those that tend to deflate at low surface tension. In the absence of surfactant, the surface tension at the airliquid interface would remain constant, and the transalveolar pressure needed to keep it at that volume would be higher when alveolar volumes are lower. Therefore, higher transalveolar pressure would be necessary to produce a given increase in alveolar volume at lower lung volumes than at higher lung volumes. Surfactant stabilizes the inflation of alveoli because it allows the surface tension to increase as the alveoli become larger. As a result, the transalveolar pressure nec essary to keep an alveolus inflated increases as lung volume and transpulmonary pressure increases, and it decreases as lung volume decreases. In the presence of surfactant, surface tension is increased at high lung volume and decreased at low lung volume. Surfactant is 85% to 90% lipids, predominantly phospholipids, and 10% to 15% proteins (Table 21. These lipids are important in formation of the monolayer on the alveolarair interface, and phosphatidylglycerol is important in the spreading of surfactant over a large surface area. Surfactant protein A is involved in the regulation of surfactant turn over, in immunoregulation within the lungs, and in the formation of tubular myelin. Surfactant is secreted into the airway through exocytosis of the lamellar body by constitutive and regulated mecha nisms. Numerous agents, including adrenergic agonists, activators of protein kinase C, leukotrienes, and purinergic agonists, stimulate the exocytosis of surfactant. Surfactant is readily inactivated by hypoxia, infection, and edema fluid, which results in a decrease in lung compliance. In addition to surfactant, another mechanism, interde pendence, contributes to stability of the alveoli. Alveoli, except for those on the pleural surface, are surrounded by other alveoli. The tendency of one alveolus to collapse is opposed by the traction exerted by the surrounding alveoli. Thus collapse of a single alveolus causes stretching and distortion of the surrounding alveoli, which in turn are connected to other alveoli. Small openings (pores of Kohn) in the alveolar walls connect adjacent alveoli, whereas the canals of Lambert connect the terminal airways to adja cent alveoli. The pores of Kohn and the canals of Lambert provide collateral ventilation and prevent alveolar collapse (atelectasis). The pressure across the respiratory system is 0 at points of no airflow (end inspiration and end exhalation). Pressure gradients in the respiratory system are created by the active contraction and subsequent relaxation of the muscles of respiration. Elastic recoil is lost in patients with emphysema, and this loss is associated with an increase in lung compliance, whereas in diseases associated with pulmonary fibrosis, lung compliance is decreased. The surface tensionreducing and antisticking properties of surfactant increase lung compliance, decrease the work of breathing, and help stabilize alveoli of different size. Lung distensibility: the static pressurevolume curve of the lungs and its use in clinical assessment. American Physiological Society Handbook of Physiology: the Respiratory System, vol. Describe how flow limitation occurs at the equal pressure point and the role of dynamic airway compression in flow limitation. Understand how dynamic compliance is different from static compliance and its contribution to work of breathing. Patterns of Airflow There are two major patterns of gas flow in the airways- laminar and turbulent. As the flow rate increases and particularly as the airways divide, the flow stream becomes unsteady and small eddies develop. The pressure-flow characteristics of laminar flow were first described by the French physician Poiseuille and apply to both liquids and air. In straight circular tubes the flow rate (V) is defined by the following equation: Equation 22. It can be seen that driving pressure (P) is proportional to the flow rate (V); thus the greater the pressure, the greater the flow. The flow resistance (R) across a set of tubes is defined as the change in driving pressure (P) divided by the flow rate, or: Equation 22. This equation is for laminar flow and demonstrates that the radius of the tube is the most important determinant of resistance. If the radius of the tube is reduced by half, the resistance will increase 16-fold. If, however, tube length is increased twofold, the resistance will increase only twofold. Stated another way, resistance is inversely proportional to the fourth power of the radius, and it is directly proportional to the length of the tube and to the viscosity of the gas.
The neurosecretory granules are then transported down the axon to the posterior pituitary and stored in the nerve endings until released symptoms 24 generic aricept 10mg otc. The osmoreceptors respond only to solutes in plasma that are effective osmoles (see Chapter 1) treatment mrsa generic aricept 5 mg fast delivery. For example medicine xalatan discount aricept 10 mg with visa, urea is an ineffective osmole when the function of osmoreceptors is considered medications via endotracheal tube purchase aricept 5 mg amex. The slope of the relationship is quite steep and accounts for the sensitivity of this system symptoms type 1 diabetes cheap aricept 10mg fast delivery. Several physiological factors can also change the set point in a given individual medications that cause pancreatitis order 5 mg aricept with amex. The mechanism responsible for the set-point shift with pregnancy is not completely understood but is likely due to hormone levels. The receptors responsible for this response are located in both the low-pressure (left atrium and large pulmonary vessels) and the high-pressure (aortic arch and carotid sinus) sides of the circulatory system. Because the low-pressure receptors are located in the high-compliance side of the circulatory system. Both groups of receptors are sensitive to stretch of the wall of the structure in which they are located. Signals from these receptors are carried in afferent fibers of the vagus and glossopharyngeal nerves to the brainstem (solitary tract nucleus of the medulla oblongata), which is part of the center that regulates heart rate and blood pressure (see also Chapter 18). Alterations in blood volume and pressure also affect the response to changes in body fluid osmolality. With a decrease in blood volume or pressure, the set point is shifted to lower osmolality values and the slope of the relationship is steeper. In terms of survival of the individual this means that faced with circulatory collapse, the kidneys will continue to conserve water, even though by doing so they reduce the osmolality of the body fluids. To compensate for this loss of water the individual must ingest a large volume of water (polydipsia) to maintain constant body fluid osmolality. This condition is called central diabetes insipidus or pituitary diabetes insipidus. In addition, their urine is more hyperosmotic than expected based on the low body-fluid osmolality. As a consequence, when water ingestion is restricted, these individuals cannot maximally concentrate their urine. This results in an increase in urea reabsorption and an increase in the osmolality of the medullary interstitial fluid, which as described below is needed for maximal urine concentration. Increasing the osmolality of the interstitial fluid of the renal medulla also increases the permeability of the inner medullary collecting duct to urea. Stimulation of thick ascending limb NaCl transport may help maintain the hyperosmotic medullary interstitium that is necessary for absorption of water from the medullary portion of the collecting duct (see below). When body fluid osmolality is increased or the blood volume or pressure is reduced, the individual perceives thirst. An increase in plasma osmolality of only 2% to 3% produces a strong desire to drink, whereas decreases in blood volume and pressure in the range of 10% to 15% are required to produce the same response. Similarly there is a genetically determined threshold for triggering the sensation of thirst. These individuals cannot maximally concentrate their urine and consequently have polyuria and polydipsia. This clinical entity is termed nephrogenic diabetes insipidus to distinguish it from central diabetes insipidus. Nephrogenic diabetes insipidus can result from a number of systemic disorders and more rarely occurs as a result of inherited disorders. Since the gene for the V2 receptor is located on the X chromosome, these inherited forms of nephrogenic diabetes insipidus are X-linked. Thus mutations in both alleles are required to produce nephrogenic diabetes insipidus. In the autosomal dominant form, the defective monomers can form tetramers with normal monomers as well as defective monomers. However, tetramers containing defective monomers are unable to traffic to the apical membrane. Recently, individuals have been found that have activating (gain-of-function) mutations in the V2 receptor gene. This new clinical entity has been termed nephrogenic syndrome of inappropriate antidiuresis. The sensation of thirst is satisfied by the act of drinking, even before sufficient water is absorbed from the gastrointestinal tract to correct the plasma osmolality. It is interesting to note that cold water is more effective in reducing the thirst sensation. Oropharyngeal and upper gastrointestinal receptors appear to be involved in this response. However, relief of the thirst sensation via these receptors is short lived, and thirst is only completely satisfied when the plasma osmolality or blood volume or pressure is corrected. However, most of the time fluid intake is dictated by cultural factors and social situations. In this situation, maintaining normal body fluid osmolality relies solely on the ability of the kidneys to excrete water. How the kidney accomplishes this is discussed in detail in the following sections of this chapter. Renal Mechanisms for Dilution and Concentration of Urine As already noted, water excretion is regulated separately from solute excretion. For this to occur the kidneys must be able to excrete urine that is either hypoosmotic or hyperosmotic with respect to body fluids. This ability to excrete urine of varying osmolality in turn requires that solute be separated from water at some point along the nephron. As discussed in Chapter 34, reabsorption of solute in the proximal tubule results in reabsorption of a proportional amount of water. Moreover, this proportionality between proximal tubule water and solute reabsorption occurs regardless of whether the kidneys excrete dilute or concentrated urine. Thus the proximal tubule reabsorbs a large portion of the filtered amount of solute and water, but it does not produce dilute or concentrated tubular fluid. The loop of Henle, in particular the thick ascending limb, is the major site where solute and water are separated. Thus excretion of both dilute and concentrated urine requires normal function of the loop of Henle. The nephron must simply reabsorb solute from the tubular fluid and not allow water reabsorption to also occur. Excretion of hyperosmotic urine is more complex and thus more difficult to understand. This mechanism is responsible for the polydipsia seen in response to the polyuria of both central and nephrogenic diabetes insipidus. Most individuals ingest water/beverages even in the absence of the thirst sensation. Normally the kidneys are able to excrete this excess water because they can excrete up to 18 L/day of urine. An example of how water intake can exceed the capacity of the kidneys to excrete water is long-distance running. A study of participants in the Boston Marathon found that 13% of the runners developed hyponatremia during the course of the race. Because over the course of the race they ingested (and generated through metabolism) more water than their kidneys were able to excrete or was lost by sweating, hyponatremia developed. In some racers the hyponatremia was severe enough to elicit the neurological symptoms described previously. For example, with maximally dilute urine (Uosm = 50 mOsm/kg H2O), the maximum urine output of 18 L/day will be achieved only if the solute excretion rate is 900 mmol/day: Uosm = Solute excretion Volume excreted 50 mOsm/kg H2O = 900 mmol/18 L If solute excretion is reduced, as commonly occurs in the elderly with reduced food intake, the maximum urine output will decrease. For example, if solute excretion is only 400 mmol/day, a maximum urine output (at Uosm = 50 mOsm/kg H2O) of only 8 L/day can be achieved. Thus individuals with reduced food intake have a reduced capacity to excrete water. Because water movement is passive, driven by an osmotic gradient, the kidney must generate a hyperosmotic compartment into which water is reabsorbed, without solute, osmotically from the tubular fluid. The hyperosmotic compartment in the kidney that serves this function is the interstitium of the renal medulla. Once established, this hyperosmotic compartment drives water reabsorption from the collecting duct and thereby concentrates urine. Establishment and maintenance of the hyperosmotic medullary interstitium has been a subject of study for more than 50 years. Despite this intense study, the most accepted model for how the medullary osmotic gradient is established, especially within the inner medulla, is incomplete and not consistent with more recent experimental findings regarding the transport properties of the nephron segments in this region of the kidney. With the caveat that the current model needs refinement, it is presented here because it embodies some fundamental concepts that underlie the process. In the current model the medullary interstitial osmotic gradient is established by a process termed countercurrent multiplication. This decreases the osmolality in the tubular fluid and raises the osmolality of the interstitium at this point. Thus at any point along the loop of Henle the fluid in the ascending limb has an osmolality less than fluid in the adjacent descending limb. Because of the countercurrent flow of tubular fluid in the descending (fluid flowing into the medulla) and ascending (fluid flow out of the medulla) limbs, this single effect could be multiplied, resulting in an osmotic gradient within the medullary interstitium, where the tip of the papilla has an osmolality of 1200 mOsm/kg H2O compared to 300 mOsm/kg H2O at the corticomedullary junction. Urea accumulates in the medullary interstitium (up to 600 mmol/L), which allows the kidneys to excrete urine with the same high urea concentration. Fluid entering the descending thin limb of the loop of Henle from the proximal tubule is isosmotic with respect to plasma. This reflects the essentially isosmotic nature of solute and water reabsorption in the proximal tubule (see Chapter 34). Most of this water is reabsorbed in the outer medulla, thereby limiting the amount of water added to the deepest part of the inner medullary interstitial space and thus preserving the hyperosmolality of this region of the medulla. In the inner medulla the terminal portion of the descending thin limb and all of the thin ascending limb is impermeable to water. This passive reabsorption of NaCl without concomitant water reabsorption begins the process of diluting the tubular fluid. The thick ascending limb of the loop of Henle is also impermeable to water and actively reabsorbs NaCl from the tubular fluid and thereby dilutes it further (see Chapter 34). Dilution occurs to such a degree that this segment is often referred to as the diluting segment of the kidney. The distal tubule and cortical portion of the collecting duct actively reabsorb NaCl. Under this condition, fluid leaving the cortical portion of the collecting duct is hypoosmotic with respect to plasma. The urine has an osmolality as low as approximately 50 mOsm/kg H2O and contains low concentrations of NaCl. An important point in understanding how a concentrated urine is produced is to recognize that while reabsorption of NaCl by the ascending thin and thick limbs of the loop of Henle dilutes the tubular fluid, the reabsorbed NaCl accumulates in the medullary interstitium and raises the osmolality of this compartment. Note also that during a water diuresis the osmolality of the medullary interstitium is reduced as a result of increased vasa recta blood flow and entry of some urea into the medullary collecting duct. This is thought to maintain the medullary interstitial gradient at a time when water is being added to this compartment from the medullary collecting duct, which would tend to dissipate the gradient. Because of NaCl reabsorption by the ascending limb of the loop of Henle, the fluid reaching the collecting duct is hypoosmotic with respect to the surrounding interstitial fluid. This diffusion of water out of the lumen of the collecting duct begins the process of urine concentration. The maximum osmolality the fluid in the distal tubule and cortical portion of the collecting duct can attain is approximately 290 mOsm/kg H2O. As the tubular fluid descends deeper into the medulla, water continues to be reabsorbed from the collecting duct, increasing the tubular fluid osmolality to 1200 mOsm/ kg H2O at the tip of the papilla. During antidiuresis, most of the water is reabsorbed in the distal tubule and cortical and outer medullary portions of the collecting duct. Thus a relatively small volume of fluid reaches the inner medullary collecting duct where it is then reabsorbed. The concentrations for NaCl and urea depicted reflect those found in the antidiuretic state. The osmotic pressure of the interstitial fluid provides the driving force for reabsorbing water from both the descending thin limb of the loop of Henle and the collecting duct. The principal solutes of the medullary interstitial fluid are NaCl and urea, but the concentration of these solutes is not uniform throughout the medulla. Establishment of the NaCl gradient is essentially complete at the transition between the outer and inner medulla. The medullary gradient for NaCl results from accumulation of NaCl reabsorbed by the nephron segments in the medulla during countercurrent multiplication. The most important segment in this regard is the ascending limb of the loop of Henle. Urea accumulation within the medullary interstitium is more complex and occurs most effectively when hyperosmotic urine is excreted.
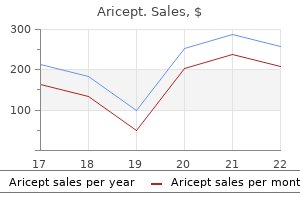
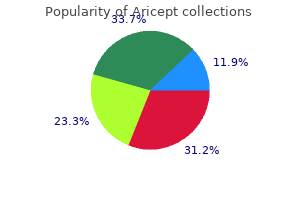
Quality 5 mg aricept. How to do Treatment of pneumonia fever | सूअरों में निमोनिया बुखार का इलाज कैसे करें.

The elevation in intracellular [Ca++] begins slightly after the action potential and peaks at approximately 20 msec symptoms 4dp5dt fet generic aricept 5 mg without a prescription. Although there is a gap (15 nm in width) between the T tubule and the terminal cisternae treatment eating disorders purchase aricept cheap online, proteins bridge this gap symptoms when quitting smoking order aricept 5 mg on line. On the basis of their appearance on electron micrographs medicine 8 letters cheap 5mg aricept overnight delivery, these bridging proteins are called feet symptoms women heart attack aricept 5mg low price. These feet are the Ca++ release channels in the membrane of the terminal cisternae that are responsible for the elevation in intracellular [Ca++] in response to the action potential medicine 48 12 purchase cheap aricept online. Calsequestrin is a low-affinity Ca++-binding protein that is present in the lumen of the terminal cisternae. It appears to bind triadin in a Ca++-dependent manner, which raises the possibility that it has a role more important than serving simply as a Ca++ buffer. Inhibition of Ca influx did not affect excitation-contraction coupling but did reduce maximal tetanic tension at high rates of electrical stimulation, which suggests that there may be some extrusion of intracellular Ca during tetany, which is compensated by Ca influx to maintain maximal tetanic tension. This pump is not unique to skeletal muscle; it is found in all cells in association with the endoplasmic reticulum. Phospholamban and sarcolipin are present in slow-twitch muscle, whereas myoregulin is present in both fast- and slow-twitch muscle. Once bound with Ca++, troponin C facilitates movement of the associated tropomyosin molecule toward the cleft of the actin filament. This movement of tropomyosin exposes myosin binding sites on the actin filament and allows a cross-bridge to form and thereby generate tension (see section "Cross-Bridge Cycling: Sarcomere Shortening"). These sites seem to be involved in controlling and enhancing the interaction between the troponin I and troponin T subunits. Although a given tropomyosin molecule extends over seven actin molecules, it is hypothesized that the strong binding of myosin to actin results in movement of an adjacent tropomyosin molecule, perhaps exposing myosin binding sites on as many as 14 actin molecules. This ability of one tropomyosin molecule to influence the movement of another may be a consequence of the close proximity of adjacent tropomyosin molecules. Such movement shortens the length of the sarcomere and thereby contracts the muscle fiber. The mechanism by which myosin produces force and shortens the sarcomere is thought to involve four basic steps that are collectively termed the cross-bridge cycle (labeled a to d in. Myosin next undergoes a conformational change termed "ratchet action" that pulls the actin filament toward the center of the sarcomere (state c). If intracellular [Ca++] is still elevated, myosin undergoes another cross-bridge cycle and produces further contraction of the muscle. The ratchet action of the cross-bridge is capable of moving the thin filament approximately 10 nm. As [Ca++] falls, Ca++ dissociates from troponin C, and the troponin-tropomyosin complex moves and blocks the myosin binding sites on the actin filament. As already noted, formation of the thick filaments involves the association of myosin molecules in a tail-totail configuration to produce a bipolar orientation. Such a bipolar orientation allows myosin to pull the actin filaments toward the center of the sarcomere during the cross-bridge cycle. The myosin molecules are also oriented in a helical array in the thick filament in such a way that cross-bridges extend toward each of the six thin filaments surrounding the thick filament. These myosin projections/cross-bridges can be seen on electron micrographs of skeletal muscle and appear to extend perpendicular from the thick filaments at rest. In the contracted state, the myosin cross-bridges slant toward the center of the sarcomere, which is consistent with the ratchet action of the myosin head. There is, however, uncertainty about how many myosin molecules contribute to the generation of force and whether both myosin heads in a given myosin molecule are involved. It has been calculated that there may be 600 myosin heads per thick filament, with a stoichiometry of 1 myosin head per 1. As a result of steric considerations, it is unlikely that all myosin heads can interact with actin, and calculations suggest that even during maximal force generation, only 20% to 40% of the myosin heads bind to actin. In isolated muscle preparations, maximum mechanical efficiency (65% efficiency) is obtained at a submaximal force of 30% maximal tension. In humans performing steady-state ergometer exercise, mechanical efficiencies range from 40% to 57%. The soleus muscle of the leg, in contrast, requires 90 msec to reach peak tension in response to an action potential, and then it relaxes slowly. The gastrocnemius muscle requires an intermediate time to reach peak tension (40 msec) because of the presence of both fasttwitch and slow-twitch muscle fibers in this muscle. Comparison of threebasic motorunit phenotypes in skeletalmuscle of extremitiesand trunk. Alterations in muscle mass and contractile phenotype in response to unloading models: role of transcriptional/ pretranslationalmechanisms. These two types of myosin isoforms have the same basic structure described previously, with two heavy chains and two pairs of light chains, although they differ in amino acid composition. It is very difficult to convert a slow-twitch muscle fiber into a fast-twitch fiber, although it can be accomplished by cross-innervation, which involves surgically interconnecting two motor neurons. Thus the motor innervation of the muscle fiber plays an important role in determining which type of myosin isoform is expressed in the muscle fiber. Further study showed that the intracellular Ca concentration in the muscle (secondary to differences in the activity pattern of the motor neuron) was an important determinant of whether the muscle fiber expressed the slow myosin isoform or the fast myosin isoform (see the section "Growth and Development"). Slow-twitch skeletal muscles are also characterized by a high oxidative capacity (see Table 12. Fast muscle, in contrast, is recruited for activities that require faster movements, more force, or both. In order to meet the demands for more force, additional motor units are recruited. In comparison with slow motor units, the fast motor units typically contain more muscle fibers (see Table 12. Thus recruitment of fast motor units can help meet the increased demands of burst activities such as weightlifting. The motor neuron in slow muscle is more easily excited than that in fast muscle, and so slow muscles are typically recruited first. The neuromuscular junction of fast muscle differs from that in slow muscle in terms of acetylcholine vesicle content, the amount of acetylcholine released, the density of nicotinic acetylcholine receptors, the acetylcholine esterase activity, and Na channel density, all of which endow the fast muscle with a higher safety factor for initiation of an action potential. During repetitive stimulation, however, the safety factor in fast muscle drops quickly (faster than that seen in slow muscle). The differential expression of troponin and tropomyosin isoforms influences the dependency of contraction on Ca++. This difference in sensitivity to Ca++ is related in part to the fact that the troponin C isoform in slow fibers has only a single low-affinity Ca++-binding site, whereas the troponin C of fast fibers has two low-affinity binding sites. Changes in the dependence of contraction on Ca++, however, are not restricted to differences in the troponin C isoforms. Thus regulation of the dependence of contraction on Ca++ is complex and involves contributions from multiple proteins on the thin filament. Increasing the Frequency of Electrical Stimulation of Skeletal Muscle Results in an Increase in the Force of Contraction. Modulation of the Force of Contraction Recruitment A simple means of increasing the force of contraction of a muscle is to recruit more muscle fibers. Because all the muscle fibers within a motor unit are activated simultaneously, a muscle recruits more muscle fibers by recruiting more motor units. Because all fibers in a motor unit are innervated by a single motor neuron, all fibers within a motor unit are of the same type. Slow-twitch motor units tend to be small (100 to 500 muscle fibers) and are innervated by an motor neuron that is easily excited (see Table 12. Fast-twitch motor units, in contrast, tend to be large (containing 1000 to 2000 muscle fibers) and are innervated by motor neurons that are more difficult to excite. The advantage of such a recruitment strategy is that the first muscle fibers recruited are those that have high resistance to fatigue. Moreover, the small size of slow-twitch motor units allows fine motor control at low levels of force. The process of increasing the force of contraction by recruiting additional motor units is termed spatial summation because forces from muscle fibers are being "summed" within a larger area of the muscle. If the muscle is stimulated a second time before it is fully relaxed, the force of contraction increases. At a high level of stimulation, intracellular [Ca++] increases and is maintained throughout the period of stimulation. At intermediate stimulus frequency, intracellular [Ca++] returns to baseline just before the next stimulus. In both cases, the increased frequency of stimulation is said to produce a fusion of twitches. The low force generation during a twitch, in comparison with that during tetany, may be due to the presence of a series elastic component in the muscle. Specifically, when the muscle is stretched a small amount shortly after initiation of the action potential, the muscle generates a twitch force that approximates the maximal tetanic force. This result, coupled with the observation that the size of the intracellular Ca++ transient during a twitch contraction is comparable with that during tetany, suggests that enough Ca++ is released into the myoplasm during a twitch to allow the actin-myosin interactions to produce maximal tension. However, the duration of the intracellular Ca++ transient during a twitch is sufficiently short that the contractile elements may not have enough time to fully stretch the series elastic components in the fiber and muscle. An increase in the duration of the intracellular Ca++ transient, as occurs with tetany, provides the muscle with sufficient time to completely stretch the series elastic component and thereby results in expression of the full contractile force of the actin-myosin interactions. As the muscle shortens, efferent output is also sent to the spindle, which thereby takes the slack out of the spindle and ensures its ability to respond to stretch at all muscle lengths. By their action, muscle spindles provide feedback to the muscle in terms of its length and thus help maintain a joint at a given angle. Slow-Twitch Muscles Exhibit Tetany at a Lower Stimulation Frequency Than Do Fast-Twitch Muscles. Thecalibrationbarfortension(ingrams)generatedduring concentration is indicated by the vertical brackets under the curves. In addition, it is likely that there are other sources of the series elastic component, such as the connective tissue and titin. The stimulus frequency needed to produce tetany depends on whether the motor unit consists of slow or fast fibers. The ability of slowtwitch muscle to tetanize at lower stimulation frequencies reflects, at least in part, the longer duration of contraction seen in slow fibers. Golgi tendon organs are located in the tendons of muscles and provide feedback regarding contraction of the muscle. The main component of the tendon organ is an elongated fascicle of collagen bundles that is in series with the muscle fibers and can respond to contractions of individual muscle fibers. A given tendon organ may attach to several fast-twitch or slow-twitch muscle fibers (or both) and sends impulses through type Ib afferent nerve fibers in response to muscle contraction. The type Ib afferent impulses enter the spinal cord, which can promote inhibition of motor neurons to the contracting (and synergistic) muscles while promoting excitation of motor neurons to antagonistic muscles. The inhibitory actions are mediated through interneurons in the cord that release an inhibitory transmitter to the motor neuron and create an inhibitory postsynaptic potential. The type Ib afferent impulses are also sent to higher centers of the brain (including the motor cortex and cerebellum). It is hypothesized that feedback from the tendon organs in response to muscle contraction may smooth the progression of muscle contraction by limiting the recruitment of additional motor units. Of interest is that the response of the tendon organ is not linearly related to force; rather, it drops off at higher levels of force, which may facilitate the recruitment of motor units at higher levels of effort. Skeletal Muscle Tone the skeletal system supports the body in an erect posture with the expenditure of relatively little energy. Nonetheless, even at rest, muscles normally exhibit some level of contractile activity. This firmness, or tone, is caused by low levels of contractile activity in some of the motor units and is driven by reflex arcs from the muscle spindles. Interruption of the reflex arc by sectioning of the sensory afferent fibers abolishes this resting muscle tone. The tone in skeletal muscle is distinct from the "tone" in smooth muscle (see Chapter 14). Modulation of Force by Reflex Arcs Stretch Reflex Skeletal muscles contain sensory fibers (muscle spindles; also called intrafusal fibers) that run parallel to the skeletal muscle fibers. The muscle spindles assess the degree of stretch of the muscle, as well as the speed of contraction. These afferent fibers in turn excite the motor neurons in the spinal cord that innervate the stretched muscle. The result is that the Energy Sources During Contraction Adenosine Triphosphate Muscle cells convert chemical energy to mechanical energy. Fatty Acids and Triglycerides Fatty acids represent an important source of energy for muscle cells during prolonged exercise. In addition, muscle cells can store triglycerides, which can be hydrolyzed when needed to produce fatty acids. For fatty acids to enter the mitochondria, however, they are converted to acylcarnitine in the cytosol and then transported into the mitochondria, where they are converted to acyl coenzyme A (CoA). Skeletal muscle fatigue during intense exercise is associated with depletion of the creatine phosphate store, although as described subsequently, this does not necessarily imply that the fatigue is caused by depletion of the creatine phosphate store. Oxygen Debt If the energy demands of exercise cannot be met by oxidative phosphorylation, an oxygen debt is incurred. After completion of exercise, respiration remains above the resting level in order to "repay" this oxygen debt.
Despite the limitations of using diffusion to describe and understand the transport of molecules across cell membranes symptoms gestational diabetes purchase 5mg aricept visa, it is also important for understanding gas exchange in the lungs (see Chapter 24) medicine 0552 aricept 5 mg generic, the movement of molecules through the cytoplasm of the cell silent treatment purchase aricept visa, and the movement of molecules between cells in the extracellular fluid treatment 4 pimples purchase aricept 5mg free shipping. For example treatment quadriceps tendonitis order generic aricept line, one of the physiological responses of skeletal muscle to exercise is the recruitment or opening of capillaries that are not perfused at rest symptoms bacterial vaginosis discount aricept 10mg fast delivery. This opening of previously closed capillaries increases capillary density and thereby reduces the diffusion distance between the capillary and the muscle fiber so that oxygen and cellular fuels. The second component (electrical potential difference) represents the energy associated with moving charged molecules. Thus for the movement of glucose across a membrane, only the concentrations of glucose inside and outside of the cell need to be considered. However, the movement of K+ across the membrane, for example, would be determined both from the K+ concentrations inside and outside of the cell and from the membrane voltage. It should be apparent that the Nernst equilibrium potential quantitates the energy in a concentration gradient and expresses that energy in millivolts. This is opposite to , and of greater magnitude than, the energy in the membrane voltage (Vm = -60 mV), which causes K+ to enter the cell. As a result, the electrochemical gradient is such that the net movement of K+ across the membrane will be out of the cell. Another way to state this is that the net driving force for K+ (Vm - E K +) is 30. It has two components: One component represents the energy in the concentration gradient for X across the membrane (chemical potential b By convention, membrane voltages are determined and reported with regard to the exterior of the cell. Positive Vm values can be observed in some excitable cells at the peak of an action potential. As shown, the glucose concentration gradient would be expected to drive glucoseintothecell. Active and Passive Transport When the net movement of a molecule across a membrane occurs in the direction predicted by the electrochemical gradient, that movement is termed passive transport. Transport that is passive is sometimes referred to as either "downhill transport" or "transport with the electrochemical gradient. Active transport is sometimes referred to as either "uphill transport" or "transport against the electrochemical gradient. When this occurs, the molecule or molecules transported against their electrochemical gradient are said to be transported by secondary active transport mechanisms. Osmosis and Osmotic Pressure the movement of water across cell membranes occurs by the process of osmosis. The movement of water is passive, with the driving force for this movement being the osmotic pressure difference across the cell membrane. Osmotic pressure is determined by the number of solute molecules dissolved in the solution. It is not dependent on such factors as the size of the molecules, their mass, or their chemical nature. Because of the presence of soluteparticles in compartment A, the concentration of water in compartment A is less than that in compartment B. Atequilibrium,thehydrostaticpressure exertedby thecolumnof water (h) stopsthe netmovementofwater from compartmentB to A. Thus at equilibrium, the hydrostatic pressure is equal and opposite to the osmotic pressure exerted by the soluteparticlesincompartmentA. Alternatively, osmotic pressure is expressed in terms of osmolarity (see the following section). Regardless of the molecule, a solution containing 1 mmol/L of the molecule therefore exerts an osmotic pressure proportional to 1 mOsm/L. For example, a 150-mmol/L solution of NaCl has an osmolarity of approximately 300 mOsm/L because each molecule of NaCl dissociates into a Na+ and a Cl- ion. Accordingly, osmolarity for any solution can be calculated as follows: Equation 1. Measurements of osmolarity are temperature dependent because the volume of the solvent varies with temperature. In contrast, osmolality, which is based on the mass of the solvent, is temperature independent. For this reason, osmolality is the preferred term for biologic systems and is used throughout this book. Because the solvent in biological solutions and bodily fluids is water, and because of the dilute nature of biological solutions and bodily solutions, osmolalities are expressed as milliosmoles per kilogram of water (mOsm/kg H2O). Tonicity the tonicity of a solution is related to the effect of the solution on the volume of a cell. A hypotonic solution causes a cell to swell, whereas a hypertonic solution causes a cell to shrink. Although related to osmolality, tonicity also accounts for the ability of the molecules in solution to cross the cell membrane. Consider two solutions: a 300-mmol/L solution of sucrose and a 300-mmol/L solution of urea. Both solutions have an osmolality of 300 mOsm/kg H2O and therefore are said to be isosmotic. When red blood cells-which for the purpose of this illustration also have an intracellular fluid osmolality of 300 mOsm/kg H2O-are placed in the two solutions, those in the sucrose solution maintain their normal volume, whereas those placed in urea swell and eventually burst. The differential effect of these solutions on Osmolarity Versus Osmolality the terms osmolarity and osmolality are frequently confused and incorrectly interchanged. Osmolarity refers to the osmotic pressure generated by the dissolved solute molecules in 1 L of solvent, whereas osmolality is the number of molecules dissolved in 1 kg of solvent. For a dilute solution, the difference between osmolarity and osmolality is c NaCl does not completely dissociate in water. In contrast, the red blood cell membrane does not contain sucrose transporters, and sucrose cannot enter the cell. To exert an osmotic pressure across a membrane, a molecule must not cross the membrane. Because the red blood cell membrane is impermeable by sucrose, it exerts an osmotic pressure equal and opposite to the osmotic pressure generated by the contents within the red blood cell (in this case, 300 mOsm/kg H2O). In contrast, urea is readily able to cross the red blood cell membrane, and it cannot exert an osmotic pressure to balance that generated by the intracellular solutes of the red blood cell. Consequently, sucrose is termed an effective osmole, whereas urea is an ineffective osmole. In contrast, = 1 for a solute that cannot cross the cell membrane (in the preceding example, sucrose). Many molecules are neither completely able nor completely unable to cross cell membranes. Specific Gravity the total concentration of all molecules in a solution can also be measured as specific gravity. Specific gravity is defined as the weight of a volume of solution divided by the weight of an equal volume of distilled water. Because biological fluids contain a number of different molecules, their specific gravities are greater than 1. Oncotic Pressure Oncotic pressure is the osmotic pressure generated by large molecules (especially proteins) in solution. The cause of this anomalous relationship between protein concentration and osmotic pressure is not completely understood, but it appears to be related to the size and shape of the protein molecule. The oncotic pressure exerted by proteins in human plasma has a normal value of approximately 26 to 28 mm Hg. Although this pressure appears to be small in relation to osmotic pressure (28 mm Hg 1. One class of these membrane proteins (membrane transport proteins or transporters) is involved in the selective and regulated transport of molecules into and out of the cell. Water movement through aquaporins is driven by an osmotic pressure difference across the membrane. Passive transport is the movement of molecules as expected from the electrochemical gradient for that molecule. Active transport is further divided into primary active and secondary active transport. Secondary active transport occurs with coupled solute carriers, for which passive movement of one or more molecules drives the active transport of other molecules. What is steady-state balance, and, with water balance as an example, what are the elements needed to achieve steady-state balance What are the volumes of the body fluid compartments, and how do they change under various conditions How do cells regulate their volume in isotonic, hypotonic, and hypertonic solutions What are the structural features of epithelial cells, how do they carry out vectorial transport, and what are the general mechanisms by which transport is regulated N ormal cellular function requires that the intracellular composition-with regard to ions, small molecules, water, pH, and a host of other substances-be maintained within a narrow range. This is accomplished by the transport of many substances and water into and out of the cell via membrane transport proteins, as described in Chapter 1. In addition, each day, food and water are ingested, and waste products are excreted from the body. In a healthy individual, these processes occur without significant changes in either the volume of the body fluid compartments or their composition. The maintenance of constant volume and composition of the body fluid compartments (and their temperature in warm-blooded animals and humans) is termed homeostasis. The human body has multiple systems designed to achieve homeostasis, the details of which are explained in the various chapters of this book. In this chapter, the basic principles that underlie the maintenance of homeostasis are outlined. In addition, the volume and composition of the various body fluid compartments are defined. Concept of Steady-State Balance the human body is an "open system," which means that substances are added to the body each day and, similarly, substances are lost from the body each day. The amounts added to or lost from the body can vary widely, depending on the environment, access to food and water, disease processes, and even cultural norms. In such an open system, homeostasis occurs through the process of steady-state balance. To illustrate the concept of steady-state balance, consider a river on which a dam is built to create a synthetic lake. At the same time, water is lost through the spillways of the dam and by the process of evaporation. Because the addition of water is not easily controlled and the loss by evaporation cannot be controlled, the only way to maintain a constant level of the lake is to regulate the amount that is lost through the spillways. To understand steady-state balance as it applies to the human body, the following key concepts are important. There must be a "set point" so that deviations from this baseline can be monitored. The sensor or sensors that monitor deviations from the set point must generate "effector signals" that can lead to changes in either input or output, or both, to maintain the desired set point. Although transient periods of imbalance can be tolerated, prolonged states of positive or negative balance are generally incompatible with life. For example, the amount of water lost through the lungs depends on the humidity of the air and the rate of respiration. Similarly, the amount of water lost as sweat varies according to ambient temperature and physical activity. Finally, water loss via the gastrointestinal tract can increase from a normal level of 100 to 200 mL/day to many liters with acute diarrhea. Of these inputs and outputs, the only two that can be regulated are increased ingestion of water in response to thirst and alterations in urine output by the kidneys (see Chapter 35). Cells within the hypothalamus of the brain monitor body fluid osmolality for deviations from the set point (normal range: 280-295 mOsm/kg H2O). The other is hormonal (antidiuretic hormone, also called arginine vasopressin), which regulates the amount of water excreted by the kidneys. With appropriate responses to these two signals, water input, water output, or both are adjusted to maintain balance and thereby keep body fluid osmolality at the set point. Volumes and Composition of Body Fluid Compartments Unicellular organisms maintain their volume and composition through exchanges with the environment they inhabit. The billions of cells that constitute the human body must maintain their volume and composition as well, but their task is much more difficult. This challenge, as well as its solution, was first articulated by the French physiologist Claude Bernard (1813-1878). He recognized that although cells within the body cannot maintain their volume and composition through exchanges with the environment, they can do so through exchanges with the fluid environment that surrounds them. He also recognized that the organ systems of the body are designed and function to maintain a constant milieu interieur or a "constant internal environment. The cardiovascular system delivers nutrients to and removes waste products from the cells and tissues and keeps the extracellular fluid well mixed. Finally, the nervous and endocrine systems provide regulation and integration of these important functions. To provide background for the study of all organ systems, this chapter presents an overview of the normal volume and composition of the body fluid compartments and describes how cells maintain their intracellular composition and volume. Included is a presentation on how cells generate and maintain a membrane potential, which is fundamental for understanding the function of excitable cells. Finally, because epithelial cells are so central to the process of regulating the volume and composition of the body fluids, the principles of solute and water transport by epithelial cells are also reviewed. Because the water content of adipose tissue is lower than that of other tissue, increased amounts of adipose tissue reduce the fraction of water in the total body weight. In some pathological conditions, additional fluid may accumulate in what is referred to as a third space.

















